




Abstract
Dimensions and shapes of fishing hooks - and their snoods- has been largely neglected from studies focused on abandoned, lost or otherwise discarded fishing gear (ALDFG) derived from angling activities in maritime areas of the globe, particularly in coastal northern Argentina. There, fieldwork was performed in a popular Argentinean coastal angling site placed within the World Reserve of Biosphere with the focus of better understanding the characteristics and the potential origin of fishhooks as an important component of ALDFG originated in recreational fishing activities. Almost 200 fishing hooks were retrieved in the context of a research and outreach conservation campaign conducted in an important recreational fishing location in south-eastern Buenos Aires province, in northern Argentine Patagonia. By applying an ensemble learning method for classification, assisted by information gathered from recreational fishermen, we were able to predict the source of overall single limerick hooks from unknown target species, estimating these fishhooks to comprise four fish-target species. These results have the potential of improving local management plans, by identifying which are the main fisheries contributing to ALDFG, hence allowing targeted campaigns to improve fishing practices.
Keywords:Recreational fisheries; Fishhooks; Abandoned, lost or otherwise discarded fishing gear; Expert consultation; Ensemble learning method; Improvement of local management plans and fishing practices
Introduction
Information regarding the sources and extent of abandoned, lost or otherwise discarded fishing gear (ALDFG) [1] like marine recreational fishhooks is particularly elusive in the tip of South America, mainly in Argentina which includes a maritime coastline that serves as a ground for a broad and diverse arrange of recreational (and artisanal) multispecies fisheries [2-4]. Like other areas of the world, local anglers capture the fish using a rod, reel, line and hook with bait or lure (often known as hook-and-line tackle) [5]. There, in northern Patagonia, the coasts of the Buenos Aires Province (1,949 km of coastline equaling ca. 30% of the entire national maritime coastline) encompass a wide variety of marine recreational angling sites that have being extensively used by anglers since the 1960s particularly in its southern section [6]. However, other northerly coastal areas are intensively used as marine angling sites like the Mar Chiquita coastal lagoon [2,7]. The area is a shallow estuarine system of 46 km2 that functions as an important nursery and feeding ground for almost 30 species of marine fish species (including both benthic and pelagic fishes) [8]. Many of these species are frequently targeted by recreational fishing activities [2,7]. The most important game species for recreational fishing due to their size and the top quality of their flesh in the area includes the Brazilian flounder (Paralichthys orbignyanus), the Black drum (Pogonias cromis) and to some extent the Silverside (Odontesthes argentinensis) [2,8]. These species are permanent year-round residents in the Mar Chiquita lagoon [8,9]. However, these game species fall within what is locally known as ´variado costero´- a group of over 30 species of both pelagic and demersal fish being exploited throughout the year by a diverse array of non-selective coastal fishing gear [10]. The area is an estuarine environment with high biological richness and diversity, considered a wildlife shelter of regional relevance [11,12]. Given its unique environmental features, it was declared a World Biosphere Reserve by MAB-UNESCO - under the name of Parque Atlántico Mar Chiquito - in 1996 and a Provincial Reserve in 1999.
Marine debris in Mar Chiquita coastal lagoon derived from recreational fishing activities has been identified amongst the most abundant superficial (non-buried) litter on sandy beaches [13-15]. Moreover, the impact of ALDFG derived from recreational fishing activities in the target area has been evidenced on entanglements in and ingestion of fish hooks and monofilament lines in seabirds like gulls, albatrosses and petrels [16,17], and other avifauna using coastal environments like raptors [18] and flamingos [19]. Regardless, no study focused on the issue of ALDFG derived from angling activities has yet addressed fishhooks and their snoods, including descriptions of dimensions and shapes at national scale. This may be due to the difficulty in sampling debris like fishhooks (along with lines) as these pose challenges for its detection given their small size and to same extent coloration, and the fact that they may be entrapped by sand and often altered by chemical and mechanical action [20]. Baseline studies of ALDFG like recreational fishhooks have the potential of improving local management plans, by identifying which are the main fisheries contributing to ALDFG, hence allowing targeted campaigns to improve fishing practices. From the fish perspective, data from hooks combined with data from damaged fish may improve our understanding about the welfare and mortality of released fish and the time needed to remove hooks and the air exposure time of the fish [21,22]. From the top predator standpoint, studies based on hooks measurements and shapes may assist biologists and managers to better understand the provenance of this fishing gear entangled or ingested by seabirds and other marine upper vertebrates, including seabirds [16,17,23].
As part of an ongoing, long-term socio-ecological study on
fisherman perception and opinions about their fishing practices
and their impacts on natural local resources at the local MAB
Biosphere Reserve [24-26] and threats posed by marine debris
derived from recreational activities in coastal settings including
this area [14,15,27], the main goals of this study were to
a) characterize the dimensions and shapes of a specific
sub-category of ALDFG represented by recreational fishing hooks
and snood configuration
b) identify the potential source (in terms of fishery and
target species) of recreational fishing hooks found in Mar Chiquita
coastal lagoon.

Methods
Study area, hook sample collection and processing
In October 2017, in the context of a research project and biodiversity conservation campaign aimed at improving recreational fishery practices and responsible disposal of fishing gear [15,24,25,27] the authors, in collaboration with anglers and visitors retrieved different components of derelict fishing gear from the sandy beach environment used in this study. Samples were inspected on the day of the survey, the presence of hooks (with or without bait and/or snood) was recorded, and placed in plastic bags for further analysis. All collected hooks were cleaned, measured and classified based on the number of bends, shape, presence of snoods, among other measurements collected as described below. The terminology for describing the features of a fishhook, which encompass the eye, shank, bend, point, barb, gape, and bite follows that described in Thomas et al. [28] and Mituhasi & Hall [29]. Recognizing that differences may arise among manufactures, fishers and researchers as to which linear and angular measurements determine hook shape and size, in this study we follow the definitions and linear measurements proposed by Thomas et al. [28]. Thus, the length of the shank - the ´leg´ of a hook - extended from the bend up to base of the eye, the eye being the portion where the (monofilament) line is attached to a hook, whereas the bend is the curved section of a hook. The length of the eye stretched from the base to the apex of the eye. The total length of the hook extended from the apex of the eye down to the base of the bend. The gape is the shortest distance between point and shank and is measured as the distance between the point and the hook shank; the point being the tip of the hook that goes into the body of the fish. The bite is the distance from the apex of the bend to its intersection with the gape [28]. The frontal length of the hook - sometimes referred to as bill - extended from the base of the bend to the point. In addition, we also quantified the hook width, this was measured as the distance between the frontal length and the length of the shank [30]. The absolute size for each hook was estimated by means of multiplying total hook length times hook width, thus this metric is expressed as mm2. All measurements were obtained aided with a digital calliper (precision 0.01 mm). All measurements were obtained by the lead author. Sampled fishhooks were described in terms of nine hook (total length of the hook, frontal length of the hook, diameter of the hook, width of the hook, length of the shank, bite, gape, length of the eye and surface) and three snood (diameter, color and the direction of the twist of the snood), binary and multistate features (Table 1). Based on the number of bends, hooks were further divided into single hooks (one bended hook forged at the top to make one eye), double hooks (two separate bended hooks forged at the top make one eye) and treble hooks (three separate bended hooks forged at the top to make one eye). Regarding their shape, single hooks were further classified into round bend hooks or ´J´ shaped hooks (showing a perfect round bend), limerick hooks (showing a sharp angle in front of the bend) and circle hooks (showing a rounded shape and its point is oriented perpendicular to the shank) [28,31]. Based on the alignment of the hook point with regards to the shank, we further divided sampled hooks into kirbed (point bent towards the right of the shank), reversed (point bent towards the left of the shank) and straight (point remains straight to the shank). Based on the design of the hook eye, sampled hooks were further classified into ringed, open, needle and flattened. Whereas based on the angle the eye makes with the shank, hooks were additionally divided into turned up (the eye is turned up from the shank), turned down (the eye is turned down from the shank) and straight (the eye is parallel to the hook bend). Finally, sampled hooks were further classified based on the type of barb in beak, knife edge, Dublin and hollow [28]. For those hooks with a snood (i.e. the knot used for connecting monofilament line to the hook), further information included the direction of the twist of the snood, which was divided into S- (the twisted line follows the direction of the central portion of the letter ´S´) and Z-twist (the twisted line follows the central portion of the letter ´Z´) [32]. Based on the color coding, snoods were further into light (encompassing colors such as white, blue, red, orange, and yellow, as well as transparent) and dark (including colors like brown, green, grey, and black) [33]. The data on white and transparent lines is presented separately for comparison purposes. Finally, we measured the diameter of the snood aided with a digital calliper (precision 0.01 mm). In this study, all snoods were comprised of plastic monofilament lines.

Expert consultation
The approach described in previous sections was supplemented with information gathered through expert consultation at six fishing tackle warehouses established within the village of Mar Chiquita and adjacent coastal areas like the village of Santa Clara del Mar and the northern section of Mar del Plata city (located 15 km and 36 km to the south of Mar Chiquita, respectively). Fishing tackle warehouses located in Santa Clara del Mar and the northern portion of Mar del Plata were chosen as these sites are on the way to Mar Chiquita. These fishing tackle warehouses have been open to trade in the area since 1970s. The information gained by traditional or non-formal local resource users like fishers from long-term use of a specific area or fishery – termed ‘fisher ecological knowledge’ - is known for being a method for collection fisheries data [34,35]. Two of the authors (MMH and FZ) performed in-person collection of data, at the two fishing tackle warehouses within the village of Mar Chiquita, regarding target species, seasonality, angling site within the study area (lagoon vs. open ocean) and technical description of gear configuration based on a hook sampler built using hooks gathered during samplings (Figure 1). Secondly, brand new hooks, some of them with snoods were purchased from all fishing tackle warehouses with a focus on main local target species like the Brazilian flounder, the Black drum and the Silverside along with species pooled as ‘variado costero’. At least three hooks with snoods when possible were gathered per game species and per fishing tackle warehouse. The Brazilian flounder, the Black drum, the Silverside and the ´variado costero´ were captured with different fishing tackle configuration, particularly hooks size and shape [2,3]. Thus, our criterion was that fishing tackle sources are based depending on the target or game species. Hook and snood binary and multistate features and measurements previously commented were also retrieved from these reference gear by the lead author.
Data analysis
The overall data set included 258 hooks. Of these, 198 hooks of unknown target species (hereinafter referred as to UTS-series) - including those used on the hook sample described above - were retrieved from the environment during samplings. Some 94 hooks from this UTS-series showed unknown or missing character states due to incomplete and/or corroded features, which accounted for 3.0% of the 3,096 possible entries for 12 characters across all gear items (Table 1). To avoid the issue of missing characters, those missing values were removed from further analysis [23]. Thus, robust UTS-series data further used in the analysis included 103 hooks with 12 characters. The other 61 reference target species gear (hereinafter referred as to RTS-series) were provided under expert consultation and purchased from local fishing tackle warehouses (Table 1). Some of this reference gears were accomplished without any snood at all, thus hook-snood data is limited in this series (Table 2). The RTS-series data included 31 hooks with 12 characters and 30 hooks with nine characters (i.e. those missing three hook-snood combinations). To determine the relationship between different hook measurements, Pearson correlation was used for each of the two variable sets (UTS and RTS-series) separately. Linear models (LMs) were used to compare the RTS-series hook measurements among target species. The response variables included total length, bite, gape, frontal length, eye length, shank, width, diameter, and surface. In all models, species were treated as a categorical variable with four levels: Black drum, Brazilian flounder, Silverside, and ‘variado costero’. To identify differences between fish species pairs, multiple comparison tests (Tukey contrasts) were conducted using the multicopy package [36]. Model assumptions were checked using DHARMa diagnostic plots from the DHARMa package [37]. To test categorical hook measurements from the RTS-series hooks among species, the Fisher Exact Test [38] was applied. Eye angle (categorized in four levels: turned up, turned down, turn up, and straight), barb shape (three levels: needle, knife edge, and beak), and point curve (three levels: straight, reversed, and kirbed) were compared across species. Pairwise species differences were assessed using the Bonferroni correction for p-values using the report tools package [39]. To compare all continuous RTS-series hook measurements among target species, pairwise multiple comparisons were conducted using Permutational Multivariate Analysis of Variance (PERMANOVA), based on Bray– Curtis distances and 999 permutations. All variables included in the PERMANOVA analysis met the assumption of homogeneity. This analysis was performed using the vegan R package [40]. Additionally, a Principal Component Analysis (PCA) was carried out to visually corroborate the results obtained from the PERMANOVA analysis. To build a model for predicting the target species based on hook measurements, a Random Forest (RF) algorithm was used [41]. The variables used to construct the model included total length, bite, gape, frontal length, eye length, shank, width, diameter, surface, eye angle, barb shape, and point curve. A total of 500 trees were built. Models with 128 [42] and 1,000 trees were also tested, but accuracy was lower or remained similar (Figure 2). Each tree within the RF was generated through recursive partitioning of the data, using the best splitting variable from a vector m randomly selected from p to partition the data at each node of the bth tree, where m was of length 2 (considering the high correlation among continuous variables and to avoid overfitting). Vectors of length 3, 9 and 12 were also tested but resulted in higher error rates (Figure 2). To validate the model, a leave-one-out cross-validation (LOOCV) procedure was employed. Out-of-bag (OOB) error estimates were examined from the LOOCV to assess prediction accuracy [41,43]. Confusion matrices were generated from OOB classifications, providing estimates of generalization error. The kappa statistic [44] was also used to measure classification performance by indicating the proportion of classification agreement beyond what would be expected by chance. Variable importance was assessed to rank the contribution of each variable to classification accuracy. The two typical measures of variable importance were calculated for the RF: mean decrease in accuracy and mean decrease in the Gini Importance Index [41]. Finally, the final RF was then used to make predictions based on the UTS-series hook measurements (i.e. single limerick hooks) to the following equation:
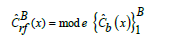
where ˆCb(x) is the classification prediction of the both tree in the ensemble of B = 500 trees, and CˆBrf (x) is the prediction of the RF. Seven single limerick hooks from the total robust UTS-series (n = 71) were excluded from the prediction, as only three single hooks contained a kriged curve of the eye, another two hooks showed a flattened hook eye, another single hook contained a Dublin barb, and another single hook contained a looped hook eye. Overall, these hooks were not used to train the algorithm. The RF was built using the random Forest [45] and caret [46] packages. To observe the grouping of the predicted hooks, a PCA analysis was performed and plotted. All the analyses were carried out using R software version 4.4.1 [47]. All values are given as mean ± standard error (se) and all tests were two-tailed with a significance level of p = 0.05.


Results
Measurements and shapes of recreational hooks from the robust UTS-series
Based on the number of bends, single hooks prevailed (81.7%, n = 85) followed only by treble hooks (18.3%). In regard to their shape, the bulk of single hooks from this series included those categorized as limerick hooks (84.7%, n = 71), followed by J hooks (14.1%), and a single live bait hook (1.17%). Table 1 summarizes overall measurements and shapes of hooks from the robust UTSseries including both single (i.e. limerick and J hooks) and treble hooks. All combinations of linear measurements from hooks from this series were significantly correlated (Pearson correlation, all p < 0.001) (Figure 3). When considering single hooks from the robust UTS-series, an average limerick hook showed a mean total length estimated at 36.6 ± 11.3 mm, bearing a mean surface of 617.9 ± 333.7 mm2. Further, a typical limerick hook from this series showed a ringed eye either straight or turned down toward the shank and a straight barb ending either as a knife edge or beak (Table 1). The average snood diameter of limerick hooks from the robust UTS-series was 0.6 mm ± 0.2 mm, and based on its frequency of occurrence, light colored snoods from these limerick hooks outnumbered dark ones. White (72.8%) prevailed as the main light color code of snoods. Snoods with an S-twist prevailed as the main twist direction of a typical limerick hook stood from this series (Table 2). In regard to (single) J hooks from the robust UTS-series, the average total hook length was computed at 40.9 ± 12.0 mm, with a mean surface estimated at 742.0 ± 330.1 mm2. An average J hook from the UTS-series showed the same features as limerick hooks from this series, i.e. a ringed eye either straight or turned down toward the shank and a straight barb ending either as a knife edge or beak (Table 1). The average snood diameter of J hooks from the robust UTS-series was 0.6 ± 0.2 mm. Based on its frequency of occurrence, light-colored snoods from these limerick hooks outnumbered dark ones. White (77.0%) prevailed as the main light color code of snoods. Both snood characters, i.e. S- and Z-twist, were equally represented in the snoods of J hooks from this series (Table 2). In the case of treble hooks belonging to the robust UTS-series, the average total length was estimated at 33.4 ± 5.5 mm, bearing a mean surface computed at 456.7 ± 161.5 mm2. A typical treble hook from the robust UTS-series showed a needle eye pointing straight toward the shank and a straight barb ending as a knife edge (Table 1). The average snood diameter for treble hooks from the robust UTS-series was computed at 0.6 ± 0.1 mm. Like both types of singles hooks previously depicted, light-colored snoods from treble hooks outnumbered dark ones. White was the single light color code of snoods. Snoods with a Z-twist prevailed as the main direction of the twist of the snood of treble hooks from this series (Table 2).

Expert consultation on a hook sampler with recreational hooks from the robust UTS-series
The in-person collection of data performed at the two fishing tackle warehouses placed within the village of Mar Chiquita revealed an overall fair degree of evenness in the identification of target species, seasonality, and angling site within the study area based on hooks retrieved from the bins (Table 3). Based on fisher ecological knowledge, four groups of hooks were spatially and seasonally identified as currently being used within the study area. This includes those aimed at the Silverside, the Black drum, the Brazilian flounder and the ´variado costero´. Besides, the inperson collection of data revealed important technical aspects of each group of hooks (Table 3).

Measurements and shapes of recreational hooks from known target species
The in-person collection of data performed at the two fishing tackle warehouses placed within the village of Mar Chiquita revealed an overall fair degree of evenness in the identification of target species, seasonality, and angling sites within the study area based on hooks retrieved from the bins (Figure 1). Fisher experts advised on the existence of four groups of hooks spatially and seasonally identified as being used within the study area, including those to target Silversides, Black drums, Brazilian flounders and other species in the ‘variado costero’ pool (Table 3). When considering hooks under the RTS-series category, but without controlling target species, single hooks prevailed (88.5%, n = 54), followed solely by treble hooks (11.4%, n = 7). Single hooks from this series included those considered as limerick hooks (98.1%, n = 53); a single hook was regarded as a live bait hook (1.8%). All combinations of linear measurements from hooks from the RTS-series were significantly correlated (Pearson correlation, all p < 0.001) (Figure 4). The average total length of limerick hooks from the RTS-series was 39.5 ± 14.9 mm, bearing a mean surface of 592.5 ± 408.1 mm2. In this series, an average limerick hook elicited a ringed eye positioned either in a straight or turned down angle with the shank and a straight barb ending as a beak or knife edge (Table 1). The average snood diameter of a sub-sample (n = 19) of limerick hooks was 0.6 ± 0.1 mm. Light-colored snoods were the only ones found among these sub-sample of limerick hooks. White (68.4%) prevailed as the main light color code of snoods. Snoods with an S-twist prevailed as the main direction of the twist of the snood of a sub-sample of limerick hooks from the RTS-series (Table 2). In the case of treble hooks belonging to the RTS-series, these showed an average total length estimated at 31.0 ± 3.9 mm. An average treble hook showed a needle eye facing straight toward the shank and a straight barb ending as a knife edge (Table 1). The average snood diameter of a sub-sample (n = 4) of treble hooks from the RTS-series was computed at 0.5 ± 0.0 mm. Light colored snoods were the only ones found among these treble hooks (as per the abundance parameter). Both snood characters, i.e. S- and Z-twist, were equally represented in the sub-sample of snoods of treble hooks from this series (Table 2). When controlling target species, (single) limerick hooks used for catching Black drum showed an average total length computed at 55.2 ± 9.8 mm, bearing a mean surface estimated at 951.1 ± 337.7 mm2. A typical hook used against P. cromis elicited a ringed eye placed in a turned down angle with the shank and a reversed barb ending as a knife edge (Table 4). In the case of (single) limerick hooks aimed at Silverside, an average total hook length was estimated at 21.7 ± 3.9 mm, bearing a mean surface estimated at 145.1 ± 59.5 mm2. An average hook utilized for catching O. argentinensis showed a ringed eye placed in a straight angle with the shank and straight barb ending as a knife edge. Regarding (single) limerick hooks used for catching Brazilian flounder, an average total hook length computed at 32.8 ± 6.5 mm, showing a mean surface estimated at 456.6 ± 173.2 mm2. An average hook used for catching P. orbignyanus showed a ringed eye placed in a straight angle with the shank and straight barb ending as a knife edge. Finally, a typical (single) limerick hook used against species lumped within the ´variado costero´ showed an average total hook computed at 40.6 ± 7.7 mm, with a mean surface estimated at 706.5 ± 239.7 mm2. An average hook of this category showed a ringed eye placed in a turn down angle with the shank and a reversed barb ending as a beak edge (Table 4).

Predicting target species in hooks from the UTS-series
Firstly, univariate analysis showed a significant effect of the game species on dimensions and shapes of hooks from known target species (LM F > 39.2, P < 0.001 in all hook dimensions, Table 5). A post hoc analysis revealed a similar pattern across all variables, except for width and surface area in relation to the target species. Total length, bite, gape, frontal length, eye length, shank, and diameter showed significant differences among species, except for Brazilian flounder vs. ‘variado costero’: the measurements of Black drum were higher than those of Brazilian flounder and ‘variado costero’, while the latter two had higher measurements than Silverside (Figure 3). Regarding width, differences were observed across all species, while no statistical difference in surface area was found between Black drum and ‘variado costero’ (Figure 5). Categorical variables such as eye angle; hook barb and curve of the barb showed an association with the game species (Fisher exact test: all p < 0.001). Secondly (multivariate analysis), and like with the previous approach, dimensions and shapes of hooks from known target species varied depending on the target species (PERMANOVA R2 = 0.69, pseudo-F = 37.16, p < 0.001). In this case, pairwise comparisons among target species showed that hooks from the Black drum and the ‘variado costero’ (PERMANOVA pseudo-F = 6.22, p = 0.08) and hooks from the Brazilian flounder and the ‘variado costero’ (PERMANOVA pseudo-F = 6.00, p = 0.10) presented nonsignificant differences. Other pairwise comparisons resulted in significant differences (PERMANOVA Black drum-Brazilian flounder pseudo-F = 23.71, p < 0.01; PERMANOVA Black drum- Silverside pseudo-F = 109.99, p < 0.01; PERMANOVA Brazilian flounder-Silverside pseudo-F = 33.16, p < 0.01; PERMANOVA Silverside-‘variado costero’ pseudo-F = 66.00, p < 0.01). Results of principal component analysis (PCA) did reveal a clustering of hooks from known target species by game species (Figure 6A). The first two axis of this PCA explained 96% (PC1: 94% and PC2: 2%) of the variability. The value of k = 0.77 was calculated from the RF confusion matrix (Table 6), where k = 0.75 is considered indicative of an excellent classifier [48]. The total OOB error rate, which represents the ratio of misclassified target species to the total number of samples, indicated an overall prediction accuracy of 83.33% for the full RF model. Silverside demonstrated the best performance with a 0% error rate, while the other target species exhibited error rates around 20% (Table 6). The diameter was the most important variable based on the mean decrease in accuracy index (Figure 7A), while the width was the most important variable according to the Gini importance index (Figure 7B). In general, continuous measurements were the better classifiers in both indices, except for the eye angle and barb in the first index (Figure 7). The Random Forest classification model predicted overall single hooks from unknown target species (Figure 6B), resulting in 24 hooks assigned to the ‘variado costero’, 20 to the Black drum, 13 to the Silverside and seven hooks corresponding to the Brazilian flounder (Table 6).






Discussion
To our knowledge, this is the first study to report data on dimensions and shapes (and snood configuration) of a specific sub-category of ALDFG represented by recreational fishing hooks in northern Argentina. Hooks and fishing lines or their parts like snoods are seldom reported in studies addressing the issue of ALDFG in coastal environments, perhaps due to their small size and confounding coloration [20]. Extant regional information about this issue is spatially biased toward a single angling site in southern Buenos Aires province in Argentina [49] and in Brazilian grounds [50-52]. Globally, available studies stemmed from the Florida Keys [53], eastern Mediterranean shores [54] (Battisti et al. 2021), the Baltic Sea [55], the Nordic countries [56], Southeast Asia [57], South Korea (Lee et al. 2023), and Sydney Harbour in Australia [58]. The majority of these studies were based on underwater surveys, thus highlighting the need for further studies addressing this issue in subtidal areas. We are not aware either of any previous study assessing the potential source of recreational fishing hooks, like what was attempted for commercial longline fisheries operating in sub-Antarctic waters [23].
Hooks from unknown target species
The hooks retrieved from the coastal environment (UTS series), were largely dominated by single limerick hooks. The high occurrence of single hooks was somehow expected, as they are the most common type used by anglers along maritime shores of northern Argentine Patagonia [59-61], including freshwater environments in Argentina [62], and elsewhere around the globe [31,56]. Anglers are perhaps intentionally using limerick hooks as these are more likely to deep-hook fish than other single hooks like circle hooks [63], particularly in heavily boned fish like the Brazilian flounder and the Black drum [64]. Single hooks are the ones allowed by the local authority for angling activities taking place in maritime areas of the Buenos Aires Province [65,66]. Based on our results, the dimensions of (single) limerick hooks were in line with those of hooks being in use by anglers in Argentina [4,65] and in southern Brazil [67], as well as artisanal fishermen along the Uruguayan Atlantic coast [68]. Other features depicting this type of hooks included the fact that overall hooks were ringed (either straight or turned down toward the shank) and barbed, with knife edge barbed hooks prevailing among these hooks. This study cannot indicate the basis for the dominance of barbed hooks, but there is previous evidence highlighting the advantages of using barbed single hooks, particularly enhanced efficiency at landing fish than barbless hooks [69]. Barbed hooks are known for implying more injuries and causing higher mortality rates relative to barbless hooks for several fish taxa [70]. Besides, anglers may spend more time unhooking fish caught using barbed hooks when compared with barbless hooks [71]. These two facts may also apply for overall hooks pertaining to the RTS-series. However, there is currently no research in place aim at comparing fishing efficiency of different types of hooks in coastal angling settings in northern Argentina, including the study area. Based on our results, the average snood diameter of (single) limerick hooks from the UTS-series were computed at 0.56 mm. This result fits with extant information, as snood diameters of single hook-and-line tackle used in Argentinian coastal angling activities varies between 0.40 mm and 0.90 mm [2,59]. In particular, the range of snood diameters used by anglers targeting the Silverside and the Brazilian flounder as well as other game species lumped within the ´variado costero´ in local coastal settings like the Mar Chiquita coastal lagoon is between 0.40 and 0.50 mm [2]. Broader snood diameters are used in southern Patagonia where anglers aim for the Broadnose sevengill shark Notorynchus cepedianus (range = 0.60 and 0.90 mm, [59]). Finally, white colored snoods with an S-twist prevailed as the main color and twist direction of (single) limerick hooks´ snoods from this series respectively. This is perhaps one of the first studies reporting coloration categories of snoods in the study area and elsewhere at a national level. The dominance of white colored snoods could be related to the belief among fisherman that this type of coloration would seem to be less visible than monofilament from other color codes and, therefore, catch more fish. Other explanation may well be that white monofilament was highly selected among fisherman because is the most common color and material in use. None of these pieces of information is currently available and this will require further research. Information about the coloration of fishing tackle lines that may end up as marine debris may assist ornithologist and managers to conserve other related resources like seabirds regarding interaction with human activities through ingestion of and entanglement in marine debris derived from ALDFG in the study area and adjacent coastal areas [16,17,72], and elsewhere in northern Argentina [49,73]. Single J hooks from the UTSseries ranked far lower (as per the abundance parameter) when compared with single limerick hooks. This could be related to the fact that this type of hook is simply less used by local anglers. Very scarce information is available for single J hooks in sportfishing activities in Argentina [4,65], but also from studies on marine debris derived from ALDFG in the southern hemisphere [50-52]. This highlights the need for updated information about their occurrence in marine environments as well as details on their dimensions and shapes as those reported here. This is also true for treble hooks, whose design is known for assisting in holding and hooking fish than a single hook [28]. The occurrence of this type of hook in our study is not surprising as a sole treble hook is allowed by the local authority in Buenos Aires Province for maritime angling [66]. There, an average hook-and-line fishing tackle aimed for the Brazilian Flounder encompass a single treble hook placed at the far end of the line [2]. Likewise, there is virtually null information as per the abundance and spatial extent in the use of this type of hook in the study area of elsewhere at the national level.
Including fisherman non-formal knowledge
A recent literature review suggested that recreational fishers´ non-formal knowledge is strongly overlooked worldwide [74]. At the local level, in the last years several sociological studies have revealed local fishers´ ecological knowledge inhabiting and/or using the Mar Chiquita coastal lagoon [24-26,75]. Angler experience is known for being recognized as a factor that impacts angling efficiency [76], fish damage, and mortality [69,77], and to some extent the associated environment [25,26]. Just one of the fishing tackle warehouses provided us with detailed descriptions of the hooks manufacturer and also dimension of hooks varied on the different manufacturers and equally by the same manufacturer [78]. That is why the manufacturing factor was not included in the analyses. It is true that a Limerick scale has been applied, with some uncertainty, to express the size of hooks in game fisheries [78]. However, we are not aware of any consolidated Limerick scale classification for hooks commercialized by local warehouses nor among fishermen. Thus, we encourage all professional and amateur anglers to keep record of the types - shapes and dimensions - of hooks used against coastal fishes which may contribute to our understanding of the diversity of hooks - and the spatial extent - currently in use in coastal angling settings in northern Argentine Patagonia. It would be highly acknowledged if such data is provided along with some basic temporal (season) and spatial (angling site) specification, which will further assist researchers and managers to inform fisheries management. This is since even at the scale of the village of Mar Chiquita the opinions of local warehouses may vary regarding their understanding of certain aspects of fishing tackle derived from unknown target species. Still, it is paramount to recognize the importance of including local non-formal knowledge in recreational fishing for conservation purposes [74]. For instance, in this study, in-person collection of data revealed important complementary technical aspects of groups of hooks that may assist in our understanding of the nature of the fishing tackle (float vs. sink tackle) based on hooks found as debris in the environment.
Predicting target species in hooks from unknown origin
In this study, dimensions and shapes of hooks from known target species varied significantly depending on the game species, presenting an arrangement based on overall general size as follows: Black drum larger than Brazilian flounder and ‘variado costero’ larger than Silverside. With increasing hook sizes, progressively larger fishes are captured [79,80]. Still, larger hooks can still hook smaller fish albeit having some size selectivity towards larger fish and may cause greater damage at the hooking site of smaller fish [81]. Thus, this variation in hook size was expected as hooks used by local anglers against these game species regularly vary in overall general measurements, but it should be recognized that hooks from different sizes may be used against more than one species [2,4,65]. Based on our results, we found a lack of significant differences between hooks clusters (except for hooks from the Black drum and the ‘variado costero’ and hooks from the Brazilian flounder and the ‘variado costero’) when using pair wise comparisons. Instead, the Random Forest is an algorithm extensively being used as a classification technique in many disciplines or fields, including Biology [82,83], Ecology [84,85] and Conservation [86,87] as it can achieve high accuracy and generalization compared to other models (e.g. linear discriminant analysis) [41,88,89]. Regarding marine debris, this algorithm has been effectively applied in studies focusing on large (> 10 cm) plastic items stranded on coastal and continental areas [90-93]. Thus, the Random Forest algorithm may be considered as a good candidate for the classification of this -and other types- of marine debris because of its accuracy and clearness [92]. Here, a suitable prediction accuracy (>80%) was obtained for our data set of hooks from unknown target species along with a k value from the Random Forest confusion matrix considered as nearly as an excellent classifier [48]. In terms of predicting game species, Silverside demonstrated the best performance with a 100% accurate rate. The most likely explanation is the clearly small size of hooks (in terms of overall dimensions) used against this species when compared to others employed towards the remaining game species. Other game species exhibited accurate rates around 80%; this may be related to a relatively low overlap in the dimensions and shapes of hooks used against the Brazilian flounder and the ´variado costero´, indicating that some anglers use hooks of relatively similar dimensions against these game species, thus precluding an excellent prediction performance. Another interest finding in this study is that the Random Forest classification model predicted overall (single) limerick hooks from unknown target species. Thus, the use of a Random Forest algorithm for smallersized debris - like fishhooks - is also attainable as demonstrated in our study. In terms of hook measurements of importance for classification purposes, two continuous measurements – the diameter and the width - outstand when considering mean decrease accuracy and mean decrease Gini importance indexes, respectively. The latter indicates the purity of classification and has been extensively used along with the Random Forest approach [94,95]. Thus, further studies with a wider hook dataset are encouraged to evaluate the usefulness of these variables in classification methods under the selected approach. Based on the RF classification model, around 70% of the hooks retrieved from the environment were assigned to angling activities focus on the ‘variado costero’ and the Black drum combined. This has direct management and health implications and thus conservation and public awareness actions should be directed toward improving the signaling for responsible fishing in the study area, conducting awareness campaigns among anglers, and/or reinforcing extant regulations particularly when angling activities are directed on these game species. It is true though that the fishes listed under the ´variado costero´ fishery are targeted by an array of fleets ranging from artisanal to coastal and high-seas fleets [10], turning the extrapolation of our results to other hook-and-line fisheries occurring along the coasts of the Buenos Aires Province. Finally, the RF classification model also showed hooks assigned to angling operations aimed at the Silverside. The size of these hooks, along with the others collected from the environment and predicted by the RF model, should also be highlighted from a conservation perspective, as these hooks if abandoned in the environment may impact a range of local and regional marine wildlife species through ingestion of and/or entanglement of this type of marine debris [16,19,96,97].
Acknowledgment
The authors appreciate the statistical advice provided by Dr. Pablo Jorgensen (Argentina). The lead author appreciates the opportunity provided by the Associate Editor Dr. Julia Hampton. Initial phases of this work were funded by the Environmental Conservation Fund of Banco Galicia and the Williams Foundation (grant FOCA2016 to G.O. García) and the National Agency for the Promotion of Research, Technological Development and Innovation (grant number PICT 2015–0262 to J.P. Seco Pon; grant number PICT 2017–1761 to M. Favero).
References
- Macfadyen G, Huntington T, Cappell R (2009) Abandoned, lost or otherwise discarded fishing gear. UNEP Regional Seas Reports and Studies, No. 185, FAO Fisheries and Aquaculture Technical Paper, No. 523. UNEP/FAO, Rome, Italy.
- Pellegrino JF, Cousseau MB (2005) Sport fishing from the coast in Mar del Plata: coastal fish. (First Edition), EMDER MGP. Mar del Plata, Argentina p. 72.
- Irigoyen AJ (2018) Fish and Sport Fishing Argentina: Sea and Patagonia. In: (1st), Sender Patagonia. Chubut, Argentina pp. 264.
- Sánchez CN, Góngora MV, Álvarez M, Parma A (2022) Artisanal fishing in Argentina: walking the country's coasts. In: (1st ), Editorial Ciudad, Buenos Aires, Argentina pp. 234.
- Huddart D (2019) Recreational Fishing. In: Huddart D, Stott T (eds.), Outdoor Recreation: Environmental Impacts and Management (1st), Palgrave Macmillan. New York, USA pp. 395-428.
- Guidi C, Baigún CRM, Ginter LG, Soricetti M, Guardiola RFJ, et al. (2021) Characteristics, preferences and perceptions of recreational fishers in northern Patagonia, Argentina. Reg Stud Mar Sci 45.
- Lucifora LO (2001) Sharks and shark fishing in Mar Chiquita. In: Iribarne O (Editor), Mar Chiquita Biosphere Reserve: Physical, Biological, and Ecological Characteristics. In: (1st), Editorial Martín. Mar del Plata, Argentina pp. 205-206.
- Cousseau MB, Díaz de Astarloa JM, Figueroa DE (2001) The ichthyofauna of Mar Chiquita Lagoon. In: Iribarne O (ed.), Mar Chiquita Biosphere Reserve: Physical, Biological and Ecological Characteristics. In: (1st), Editorial Martín. Mar del Plata, Argentina pp. 187-203.
- González CM, Díaz DAJM, Cousseau MB, Figueroa DE, Delpiani SM, et al. (2009) Fish composition in a south-western Atlantic temperate coastal lagoon: spatial–temporal variation and relationships with environmental variables. J Mar Biol Assoc UK 89: 593-604.
- Perrotta RG, Ruarte C, Carozza C (2007) Coastal fishing in Argentina. Ciencia Hoy 17: 32-43.
- Iribarne O (2001) Mar Chiquita Biosphere Reserve: Physical, biological and ecological characteristics. In: (1st), Editorial Martín. Mar del Plata, Argentina pp. 319.
- Isacch JP, Bó MS, Vega LE, Favero M, Baladrón AV, et. al. (2016) Assessing tetrapod diversity in a mosaic of environments in the southeastern Pampas ecoregion as a tool for conservation planning. Journal of the Argentine Museum of Natural Sciences 18: 211-233.
- Goodall RNP (1990) Surveys of Marine Debris on the Coasts of Argentina and Uruguay. US Marine Mammal Commission. Unpublished report.
- Becherucci ME, Rosenthal AF, Seco Pon JP (2017) Marine debris in beaches of the Southwestern Atlantic: An assessment of their abundance and mass at different spatial scales in northern coastal Argentina. Mar Pollu Bull 119: 299-306.
- Seco Pon JP, Hernandez MM, Zumpano F, Castano MV, Favero M, et al. (2023a) Assessment of marine debris on the Mar Chiquita coastal lagoon (Biosphere Reserve, MAB-UNESCO), a unique wetland in northern Argentina. OCMA 239: 106604.
- Berón MP, Favero M (2009) Mortality and injuries of Olrog’s Gull (Larus atlanticus) individuals associated with sport fishing activities in Mar Chiquita coastal lagoon, Buenos Aires Province. El Hornero 24: 99-102.
- Berón MP, Seco Pon JP (2021) Fishing-gear related injuries and mortality of seabirds in coastal areas of south- western Atlantic, Argentina. Mar Ornithology 49: 321-327.
- Seco Pon JP, Bó MS, Block C, Galván FE, García GO (2018) Chimango caracara (Milvago chimango) entangled in fishing tackle in southeastern Buenos Aires Province, Argentina. Ornitol Neotrop 29: 271-274.
- Berón MP (2019) Austral Flamingo Phoenicopterus chilensis sustaining an injury derived from recreational fishing gear. IJSR 8: 382-386.
- Battisti C, Gallitelli L, Vanadia S, Scalici M (2023) General macro-litter as a proxy for fishing lines, hooks and nets entrapping beach-nesting birds: Implications for clean-ups. Mar Poll Bull 186: 114502.
- Arlinghaus R, Klefoth T, Kobler A, Cooke SJ (2008) Size selectivity, injury, handling time, and determinants of initial hooking mortality in recreational angling for northern pike: the influence of type and size of bait. N Am J Fish Manag 28(1): 123-134.
- Stålhammar M, Linderfalk R, Brönmark C, Arlinghaus R, Nilsson PA (2012) The impact of catch-and-release on the foraging behaviour of pike (Esox lucius) when released alone or into groups. Fish Res 125-126: 51-56.
- Ridley C, Harrison NM, Phillips RA, Pugh PJA (2010) Identifying the origins of fishing gear ingested by seabirds: a novel multivariate approach. Aquat Conserv: Mar Freshw Ecosyst 20: 621-631.
- Cabral V, García GO, Zulaica L (2022) The Mar Chiquito Atlantic Park Biosphere Reserve: a socio-environmental assessment from the perspective of the local community. Socio territorial Studies - Journal of Geography 31: 103.
- García GO, Cabral VN, Zumpano F, Gorostegui VA (2022) Anglers’ perception and attitudes towards angling related marine litter and a conservation program in Argentina. OCMA 230.
- Zumpano F, Copello S, Cabral V, Favero M, García GO (2024) Interaction between coastal birds and recreational fisheries in Argentine: Incorporating the social dimension to improve effectiveness of conservation and management measures in a key estuarine reserve. Mar Pol 167.
- García GO, Gorostegui VA, Zumpano F, Hernández M, Friedman I, et al. (2021) Conservation approach in a coastal reserve in Argentina to promote the responsible disposal of litter derived from recreational fisheries. OCMA 214.
- Thomas SN, Edappazham G, Meenakumari B, Ashraf PM (2007) Fishing hooks: A review. Fish Tech 44: 1-16.
- Mituhasi T, Hall M (2011) Hooks used in artisanal longline fisheries of the Eastern Pacific Ocean. (First Edition), Inter-American Tropical Tuna Commission, California, USA p. 28.
- Grixti D, Conron SD, Jones PL (2007) The effect of hook/bait size and angling technique on the hooking location and the catch of recreationally caught black bream Acanthopagrus butcheri. Fish Res 84(3): 338-344.
- Serafy JE, Cooke SJ, Diaz GA, Graves JE, Hall M, et al. (2012) Circle hooks in commercial, recreational, and artisanal fisheries: research status and needs for improved conservation and management. Bull Mar Sci 88(3): 371-391.
- Radhalekshmy K, Gopalan NS (1973) Synthetic fibres for fishing gear. Fish Tech 10: 142-165.
- Verlis KM, Campbell ML, Wilson SP (2014) Marine debris is selected as nesting material by the brown booby (Sula leucogaster) within the Swain Reefs, Great Barrier Reef, Australia. Mar Poll Bull 87(1-2): 180-190.
- Hind EJ (2015) A review of the past, the present, and the future of fishers' knowledge research: a challenge to established fisheries science. ICES J Mar Sci 72: 341-358.
- Pita P, Antelo M, Hyder K, Vingada J, Villasante S (2020) The use of recreational fishers’ ecological knowledge to assess the conservation status of marine ecosystems. Front Mar Sci 7: 242.
- Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biom J 50(3): 346-363.
- Hartig F (2022) DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models_. R package version 0.4.6.
- Agresti A (2002) Categorical data analysis. In: (2nd), John Wiley & Sons, New York, USA. pp. 394.
- Rufibach K (2009) reporttools: R Functions to Generate LaTeX Tables of Descriptive Statistics. Journal of Statistical Software, Code Snippets 31.
- Oksanen J, Simpson G, Blanchet F, Kindt R, Legendre P, et al. (2024) Vegan: Community Ecology Package. R package version 2 p. 6-8.
- Breiman L (2001) Random forests. Mach Learn 45: 5-32.
- Oshiro TM, Perez PS, Baranauskas JA (2012) How many trees in a random forest? 8th International Conference Machine Learning and Data Mining in Pattern Recognition, Germany.
- Hastie T, Tibshirani R, Friedman J (2009) The Elements of Statistical Learning. In: (2nd), Springer, London, UK pp. 764.
- Cohen J (1960) A coefficient of agreement for nominal scales. Educ Psychol Meas 20(1): 37-46.
- Liaw A, Wiener M (2002) Classification and Regression by random Forest. R News 2(3): 18-22.
- Kuhn M (2008) Building Predictive Models in R Using the caret Package. J Statist Softw 28(5): 1- 26.
- R Core Team (2024) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
- Fielding A, Bell J (1997) A review of methods for the assessment of prediction errors in conservation presence/absence models. Environmental Conservation 24(1): 38-49.
- Yorio P, Marinao C, Suárez N (2014) Kelp Gulls (Larus dominicanus) killed and injured by discarded monofilament lines at a marine recreational fishery in northern Patagonia. Mar Poll Bull 85(1): 186-189.
- Farias EG, Preichardt PR, Dantas DV (2018) Influence of fishing activity over the marine debris composition close to coastal jetty. ESPR 25: 16246-16253.
- Adelir AJ, Rocha GRA, Souza TF, Pinheiro PC, Freire KDM (2016) Abandoned, lost or otherwise discarded fishing gears in rocky reefs of Southern Brazil. Brazilian J Oceanography 64(4): 427-434.
- Link J, Segal B, Casarini LM (2019) Abandoned, lost or otherwise discarded fishing gear in Brazil: A review. Perspec Ecol Conserv 17(1): 1-8.
- Chiappone M, Dienes H, Swanson DW, Miller SL (2005) Impacts of lost fishing gear on coral reef sessile invertebrates in the Florida Keys National Marine Sanctuary. Biol Conserv 121(2): 221-230.
- Battisti C, Kroha S, Kozhuharova E, De Michelis S, Fanelli G, et al. (2019) Fishing lines and fish hooks as neglected marine litter: first data on chemical composition, densities, and biological entrapment from a Mediterranean beach. ESPR 26: 1000-1007.
- Kammann U, Nogueira P, Wilhelm E, Int VI, Aust MO, et al. (2023) Abandoned, lost or otherwise discarded fishing gear (ALDFG) as part of marine litter at the seafloor of the Baltic Sea - Characterization, quantification, polymer composition and possible impact. Mar Poll Bull 194.
- Unsbo H, Granberg M, Boltenstern M, Olshammar M, Karlsson M (2022) Quantification and environmental pollution aspects of lost fishing gear in the Nordic countries. The Swedish Agency for Marine and Water Management Report number C775.
- Gajanur AR, Jaafar Z (2022) Abandoned, lost, or discarded fishing gear at urban coastlines. Mar Poll Bull 175: 113341.
- Sherow B, Gacutan J, Tait H, Johnston EL, Clark GF (2023) Land use and COVID-19 lockdowns influence debris composition and abundance in stormwater drains. STOTEN 871.
- Caille G, Cedrola P (2007) Sport fishing for the leopard shark Notorynchus cepedianus in the Deseado Estuary and the San Julián Peninsula. Patagonia Natural Foundation, Puerto Madryn, Argentina.
- Llompart FM, Colautti DC, Baigún CRM (2012) Assessment of a major shore-based marine recreational fishery in the Southwest Atlantic, Argentina. N Z J Mar Fresh Res 46(1): 57-70.
- Guidi C (2019) Analysis of recreational fishing, users, and their practices in three North Patagonian fisheries to contribute to their management. Bachelor Thesis, Universidad Nacional de Río Negro, Argentina.
- Gagne TOK, Ovitz L, Griffin LP, Brownscombe JW, Cooke SJ, et. al. (2017) Evaluating the consequences of catch and release recreational angling on golden dorado (Salminus brasiliensis) in Salta, Argentina. Fish Res 186(3): 625-633.
- Skomal GB, Chase BC, Prince ED (2002) A comparison of circle hook and straight hook performance in recreational fisheries for juvenile Atlantic bluefin tuna. Am Fish Soc Symp 30: 57-65.
- Cousseau MB, Perrotta RG (2004) Marine fishes of Argentina: biology, distribution, fishing. In: (1st), INIDEP. Mar del Plata, Argentina, pp. 193.
- Venerus LA, Cedrola PV (2017) Review of marine recreational fisheries regulations in Argentina. Mar Pol 81: 202-210.
- Ministry of Agrarian Development (MDA) (2019) Sport Fishing Regulations. Province of Buenos Aires, Buenos Aires, Argentina p. 20.
- Boaventura ICdR, Ribeiro VL, Americano AP, Taccani GDR, Ogkawara MH, et al. (2022) Impact of recreational fishing on seagull (Larus dominicanus) rescued on the coast of the São Paulo state: case report. Acta Vet Bras 16(2): 101-104.
- Ligrone A, Franco TV, Passadore C, Szephegyi MN, Carranza A (2014) Fishing strategies and spatial dynamics of artisanal fisheries in the Uruguayan Atlantic coast. Lat Am J Aquat Res 42: 1126-1135.
- Meka JM (2004) The influence of hook type, angler experience, and fish size on injury rates and the duration of capture in an Alaskan catch-and-release rainbow trout fishery. N Am J Fish Manag 24(4): 1309-1321.
- DuBois RB, Kuklinski KE (2004) Effect of hook type on mortality, trauma, and capture efficiency of wild, stream-resident trout caught by active baitfishing. N Am J Fish Manag 24(2): 617-623.
- Czarkowski TK, Kupren K, Hakuć BA, Kapusta A (2023) Fish hooks and the protection of living aquatic resources in the context of recreational catch-and-release fishing practice and fishing tourism. Water 15: 1842.
- Seco Pon JP, Álvarez V, Torres NA, Rosenthal AF, García GO (2023b) Ingestion of marine debris by Magellanic penguins (Spheniscus magellanicus) in wintering grounds of coastal Argentina. Mar Poll Bull 193.
- Canti S, González P, Suárez N, Yorio P, Marinao C (2023) Interactions between breeding gulls and monofilament lines at one of the main recreational fishing sites in Argentina. Mar Poll Bull 188: 114720.
- Löki V, Nagy J, Neményi Z, Hagyó A, Nagy A, et. al. (2023) Exploring ecological knowledge in recreational fishing for conservation purposes: A literature review. GECCO 48.
- Cabral V, Gorostegui VA, García GO (2019) Development as a frontier of conservation. Reflections on collaborative conservation in protected natural areas. Letras Verdes 26: 35-50.
- Heermann L, Emmrich M, Heynen M, Dorow M, König U, et al. (2013) Explaining recreational angling catch rates of Eurasian perch, Perca fluviatilis: the role of natural and fishing‐related environmental factors. Fish Manag Ecol 20(2-3): 187-200.
- Dunmall KM, Cooke SJ, Schreer JF, McKinley RS (2001) The effect of scented lures on the hooking injury and mortality of smallmouth bass caught by novice and experienced anglers. N Am J Fish Manag 21(1): 242-248.
- Modassir Y (2011) Fish species diversity and its potential for angling fishing in Goa - An overview. Indian J App Pure Biol 26: 291-304.
- Alós J, Cerdà M, Deudero S, Grau AM (2008) Influence of hook size and type on short-term mortality, hooking location and size selectivity in a Spanish recreational fishery. J Appl Ichthyology 24: 658-663.
- Cerdà M, Alós J, Palmer M, Grau AM, Riera F (2010) Managing recreational fisheries through gear restrictions: the case of limiting hook size in the recreational fishery from the Balearic Islands (NW Mediterranean). Fish Res 101(3): 146-155.
- Cooke SJ, Suski CD (2004) Are circle hooks an effective tool for conserving marine and freshwater recreational catch-and-release fisheries? Aquat Conserv: Mar Freshw Ecosyst 14(3): 299-326.
- Liu S, Chen Y, Wilkins D (2012) Large margin classifiers and random forests for integrated biological prediction. IJBRA 8(1-2): 38-53.
- Touw WG, Bayjanov JR, Overmars L, Backus L, Boekhorst J, et al. (2013) Data mining in the Life Sciences with Random Forest: a walk in the park or lost in the jungle? Brief Bioinform 14(3): 315-326.
- Cutler DR, Edwards Jr TC, Beard KH, Cutler A, Hess KT, et. al. (2007) Random forests for classification in ecology. Ecology 88(11): 2783-2792.
- Akselrud CIA (2024) Random forest regression models in ecology: Accounting for messy biological data and producing predictions with uncertainty. Fish Res 280: 107161.
- Mosebo FAC, Quintero GR, Lenihan CMA, Leslie TEF, Jokar AJ (2020) Machine learning for conservation planning in a changing climate. Sustainability 12: 7657.
- Madarász B, Jakab G, Szalai Z, Juhos K, Kotroczó Z, et al. (2021) Long-term effects of conservation tillage on soil erosion in Central Europe: A random forest-based approach. Soil Till Res 209.
- Fallon NG, Fielding S, Fernandes PG (2016) Classification of Southern Ocean krill and icefish echoes using random forests. ICES J Mar Sci 73: 1998-2008.
- Wang R (2022) Comparison of decision tree, random forest and linear discriminant analysis models in breast cancer prediction. J Phys: Conf Ser 2386: 012043.
- Gonçalves G, Andriolo U, Gonçalves L, Sobral P, Bessa F (2020) Quantifying marine macro litter abundance on a sandy beach using unmanned aerial systems and object-oriented machine learning methods. Remote Sens 12: 2599.
- Cortesi I, Masiero A, De Giglio M, Tucci G, Dubbini M (2021) Random forest-based river plastic detection with a handheld multispectral camera. ISPRS 43: 9-14.
- Nagy M, Istrate L, Simtinică M, Travadel S, Blanc P (2022) Automatic detection of marine litter: a general framework to leverage synthetic data. Remote Sens 14: 6102.
- Jayabhavani GN, Muthu T (2024) Floating litter detection at the estuary of Puducherry using Sentinel‐2 data and machine learning model. Reg Stud Mar Sci 77.
- Qi Y (2012) Random forest for bioinformatics. Ensemble Mach Learn: Methods App pp. 307-323.
- Han H, Guo X, Yu H (2016) Variable selection using mean decrease accuracy and mean decrease Gini based on random forest. 7th IEEE International Conference on Software Engineering and Service Science (ICSESS), China.
- Denuncio P, Mandiola MA, Salles SBP, Machado R, Ott PH, et al. (2017) Marine debris ingestion by the South American fur seal from the Southwest Atlantic Ocean. Mar Poll Bull 122(1-2): 420-425.
- Machado R (2022) Ingestion of fishing items by South American sea lion in southern Brazil: Indicating a neglected problem. Fish Res 255: 106459.















