




Greenhouse Gas Fluxes and Carbon Stock in Conserved and Degraded Mangroves
Espinosa-Fuentes María de la Luz1*, Cerón-Bretón Julia G2, Cerón-Bretón Rosa M2, Peralta Oscar1, Castro Telma1, Martínez-Arroyo Amparo1 and Hernández-Solís José Manuel1
1Department of Environmental Sciences, Institute of Atmospheric Sciences and Climate Change, National Autonomous University of Mexico, México
2Department of Chemistry, Autonomous University of Carmen, México
Submission: July 24, 2024; Published: August 09, 2024
*Corresponding author: Espinosa-Fuentes María de la Luz, Department of Environmental Sciences, Institute of Atmospheric Sciences and Climate Change, National Autonomous University of Mexico, México City 04510, Mexico, Email ID: marilu@atmosfera.unam.mx
How to cite this article: Espinosa-Fuentes M d l L, Cerón-Bretón Julia G, Cerón-Bretón Rosa M, Peralta Oscar1, Castro T, Martínez-Arroyo A, et al. Greenhouse Gas Fluxes and Carbon Stock in Conserved and Degraded Mangroves. Int J Environ Sci Nat Res. 2024; 33(5): 556375. DOI: 10.19080/IJESNR.2024.33.556375
Abstract
Mangrove degradation in tropical areas is an alarming phenomenon with significant local and global consequences for greenhouse gas (GHG) emissions and carbon storage (SCS). In this study, seasonal fluxes of carbon dioxide (CO2), methane (CH4) and nitrous oxide (N2O) from soil and SCS were evaluated in mangroves with different ecological conditions (conserved and degraded) in the western coastal sector of Campeche, Mexico. The highest CO2 fluxes (1108.5 to 1563.7mg m-2 h-1) were recorded during the dry season in the conserved mangroves. CH4 fluxes were highest in the most degraded mangroves during the wet season (6071.4 to 6675.47µg m-2 h-1). N2O fluxes did not show significant differences between treatments (p>0.05), with the highest fluxes occurring during the rainy season in the Atasta mangrove forest (713.6 ± 258.1µg m-2 h-1). The highest contribution to global warming potential among seasons and sites was CO2 (62.2 ± 34.1 Mg CO2-eq ha-1 yr-1), followed by N2O (15.7 ± 4.5 Mg CO2-eq ha-1 yr-1) and finally CH4 with the lowest CO2-eq contribution of 9 ± 5.7 Mg CO2-eq ha-1 yr-1. The conserved mangroves stored an average of 503.6 ± 96.7 Mg C ha-1 and the more degraded mangroves stored 275.1 ± 25 Mg C ha-1. The results of the present study suggest that GHG emissions and SCS are strongly influenced by site ecological conditions, seasonality and variation in some soil properties so that the ecological degradation of mangroves can increase GHG emissions and reduce the ecosystem’s capacity to store carbon.
Keywords: Tropical mangroves; GHG fluxes; Soil carbon storage
Introduction
The continuous and massive release of greenhouse gases (GHG) has led to a global rise in Earth's temperature. GHGs are emitted from fossil fuel combustion and use, industrial activities, agriculture, land use change and natural sources. The natural sources such as wetlands that are among the most carbon-rich natural ecosystems in the world [1]. Coastal wetlands such as mangroves found in tropical and subtropical regions, are unique ecosystems that play a crucial role in coastal stability. They protect against extreme weather events, provide vital habitats for numerous marine and terrestrial species, and offer a variety of ecosystem services including high rates of carbon sequestration and storage [2-5].
In recent decades, mangroves have experienced a high degradation rate due to various anthropogenic factors, such as conversion for agriculture, aquaculture, urban development, pollution, and natural resource extraction [6]. The rate of degradation results in the loss of habitats and biodiversity, as well as the disruption of ecosystem services involving global climate change [7,8].
One of the most significant impacts of mangrove degradation is its contribution to GHG emissions, particularly carbon dioxide (CO2), methane (CH4) and nitrous oxide (N2O). Mangroves are effective carbon sinks, storing large amounts of carbon in their biomass and soil. However, when degraded or destroyed, this stored carbon is released back into the atmosphere [9,10]. In addition, mangrove degradation also leads to the decomposition of accumulated organic matter in the soil, releasing more CO2 and CH4 into the atmosphere.
The emission of GHG in mangroves depends on the variation of physicochemical and environmental parameters such as the sediment grain size, the extent of flooding and temperature, the intertidal position, the redox conditions of sediments, the soil pH and salinity [11].
The dynamics of GHG emission and carbon accumulation in mangrove soils in tropical regions, like Mexico, have been scarcely studied despite carbon stocks in Mexican mangroves ranging from 79 to 647 Mg ha−1 [12]. It has been estimated the total carbon stock in degraded mangroves of the country is 349 ± 6 Tg C and emits an annual average of 0.09 ± 0.04 Tg CO2e yr−1 [13].
Our objective was to measure the variation in soil CO2, CH4, and N2O fluxes in mangroves with different conditions of ecological degradation to determine whether degraded mangrove soil is a significant source of CO2, CH4, and N2O, besides quantify the carbon stock in both well-preserved and degraded mangroves and identify the major environmental factors affecting soil GHG flux and carbon storage.
Materials and Methods
Study site description
The study area is in the south of the Gulf of Mexico, in the western sector of the coast of Campeche, in between Nuevo Campechito and Boca Nueva, on the shore of the Natural Protected Area Terminos Lagoon in Carmen Island, Campeche, Mexico (Figure 1). The diversity in species and habitats, the marine resources, the interrelation with Campeche Sound (an important fishing area in the Gulf of Mexico), make this site one of Mexico's most important lagoon systems. It is part of the National Flora and Fauna Reserve Terminos Lagoon that comprises 705,016ha of open water and associated wetlands and upland, including associated lagoons and channels, with an average depth of 4m, surrounded by 259,000ha of mangrove and marsh [14]. The region is part of the ecological complex of the coastal plain that controls the deltaic processes of the Grijalva-Usumacinta River system.

Terminos Lagoon is surrounded by mangrove forest (Rhizophora mangle, Avicennia germinans, Laguncularia racemosa, and Conocarpus erectus), covering an area as large as the open-water area of the lagoon. Therefore, mangrove constitutes a significant component of this ecosystem. The climate of the coastal plain of the Gulf of Mexico has four seasons: dry from February to April, transition in May, wet from June to September, and nortes from October to March [15]. In the Gulf of Mexico, the windy season is locally called nortes and is associated with low temperatures, northerly winds up to 100km h-1, and rainfall. The annual precipitation fluctuates between 1200 and 1650mm. The annual temperature varies between 17°C and 37°C.
We considered the dry, rainy and nortes seasons on the sampling campaigns. Field work was conducted during March and June 2019 and January 2020. The study covered five mangrove sites with different conditions of ecological degradation: Atasta (ATA), Estero Pargo (EP), Nuevo Campechito conserved (NCC) and Nuevo Campechito degraded (NCD). On the nortes and rainy seasons we also sampled at Tortuguero (TOR).
The mangrove’s ecological condition was classified as good or conserved, fair and poor. A conserved site refers to a mangrove forest that is in natural condition, with high vegetation density with heights ranging from 4 to 8m. The fair condition refers to vegetation with heights varying from 1 to 2m, with degraded mangrove or natural regeneration of mangrove. Poor condition refers to dead and degraded mangroves. The location and characteristics of each monitoring site are presented in Figure 1 & Table 1.

Greenhouse gas samplings and flux measurements
At each mangrove site, a 4 x 12m plot was established considering free access to the zone, risks, mangrove vegetation structure and distribution. Closed chambers collected CO2, CH4, and N2O from the sediment-atmosphere interface. Three chambers sampled at each site. The chambers consisted of a PVC pipe (diameter 19.5cm, length 14.5cm, volume 4.33L and area of 0.029m2). The open end of the chambers was inserted 3cm into the soil. The deployment time was set at 80min with a sampling of 20 minutes intervals. The chambers have a port which uses an injection septum for gas chromatography, and with a vacutainer needle sampled 20ml in headspace-type vials, previously evacuated in the laboratory at approximately -500mbar.
The samples were analyzed with a gas chromatographer Shimadzu model GC-2014AFsc coupled to a methanizer with flame ionization detector (FID), with a Hayesep D column of 80/100 13 ft X 1/8 in. The carrier gas was N2 5.0 UHP at 25ml min-1. The instrument operated under isothermal conditions (80° C). For the calibration curves we employed certified standard of CH4, CO2 and NO2 at 500, 6161 and 5ppmv respectively in nitrogen balance. Analytical uncertainty was ± 5%.
GHG flux was calculated with a linear regression of gas concentration in the chamber over time. Only regressions with r2 values ≥ 0.8 were used for flux calculations from all gases and all sites following Howard et al. [16]. GHG efflux rate (Fx) was calculated as:
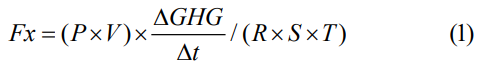
where P is the atmospheric pressure (assume the value = 1 atmosphere), V is the volume of the chamber (L), R is the ideal gas constant = 0.0820 (L*atm/K*mol) , S is the surface area covered by each chamber (m-2), T is the temperature in Kelvin degrees at the time of each measurement (K= 273 + temp in °C), and ΔGHG/Δt is the change in GHG concentration over time (Δt) based on the slope of linear regression.
Based on the average gas fluxes of the different mangrove zones, the CH4 and N2O were converted to CO2-equivalent (CO2-eq) to indicate the gas warming effect [17]. The CO2-equivalent flux was calculated using the following formula:
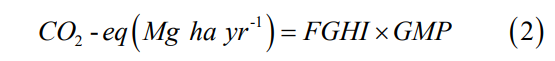
where FGHI are annual fluxes of CO2, CH4, and N2O between terrestrial and atmospheric ecosystems as a function of C and N mass, GMP is the warming effect or the conversion of CO2, CH4 and N2O emissions to CO2 equivalents as 1, 27 and 273, respectively, over a 100-year timeframe [17].
Sampling and analyses of soils
To determine the physicochemical properties of the soils, we plot three sampling points of 1m2 at each mangrove and took four soil core samples at 0-30 and 30-60cm depth using a sampler of 193.3cm3. In total 84 samples.
The samples were stored in plastic bags and transported to the laboratory. Soil samples were dried at room temperature, pulverized, and passed through a 2mm sieve. All field samples were processed following standard protocols for quantifying mangrove soils. All visible organic matter (roots, pneumatophores and leaves) were removed from the soil samples prior to further analysis. The temperature of soil (Ts) was determined in situ, using a Redox ORP 14 electrode (220mV/pH7) connected to a pH/mV/T meter PCE-228. The sediment sample was diluted with distilled water 1:5, and then the probe was inserted into the cleared solution to measure the redox potential (Eh).
Soil moisture (SM) was determined according to Etchevers et al. [18]. The pH was determined with a HANNA (Instruments electronic) model pH211. The pH meter was calibrated with two buffer solutions 7 and 4 Merck Certipur (DEU). The soil electrical conductivity (EC) was measured using a TRANS instrument model HC3010, TDS-Conductivity-Salinity (USA), with an electrode TIPB10-0400 with K = 0.9660 and calibrated with a standard solution of 1.4mS. The pH and EC were measured from aqueous extracts and verified in duplicate after 1min, followed by stirring of samples for 30s. The aqueous extracts were prepared with deionized water (>18.2 MΩcm) and CaCl2 0.01 M ACS certified (Japan) (CaCl2·2H2O, Fisher Scientific, Hampton, NH, USA) solution. The determination of soil texture was made following Bouyouco's method [19]. For bulk density (BD), an undisturbed sample of soil was taken using a steel cylinder (95.4cm3). BD was determined from the oven-dried (105°C) mass of the core and the core volume.
We analyzed Cl−, NO3 − and SO4 2− by anion exchange chromatography, applying the high-resolution liquids chromatography technique (HPLC).
Soil organic matter percentage (SOM) was quantified using the loss on ignition (LOI) method [20]. The determination of soil organic carbon percentage (SOC) was calculated using the equation proposed by Kauffman & Donato [21]:
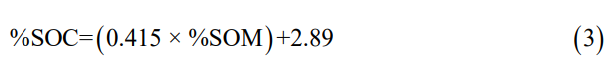
The soil carbon stock (SCS) per sampled depth interval was calculated according to Kauffman & Donato [21]:
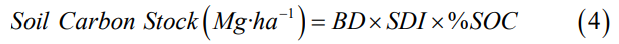
where BD is the bulk density (g·cm-3) and SDI is the soil depth interval (cm). The total SCS by site was calculated with the C stocks of both soil depth ranges (0-30 and 30-60cm).
Statistical analysis
The assumption of normality and homogeneity of variances was tested using the Shapiro–Wilk and Levene tests. The data did not have a normal distribution, so we applied the Kruskal-Wallis test. This analysis determined the differences in greenhouse gas fluxes and sediment physicochemical properties among sampling seasons and mangrove sites. If the difference was significant at p <0.05, then a post-hoc Bonferroni test was used to determine the difference. We used multivariate principal component analysis (PCA) to assess the relationships between sediment properties and GHG as well as the two ecological conditions (conserved vs degraded mangroves). Statistical analysis was conducted using SPSS Statistics v27.0 (SPSS Inc., Armonk, New York).
Results and Discussion
Seasonal variability of CO2, CH4 and N2O fluxes at the sediment-air interface and their physicochemical drivers
Based on the Kruskal-Wallis test, CO2, CH4, and N2O fluxes varied significantly between sampling periods (p< 0.05).
Carbon dioxide fluxes
CO2 flux ranged between 184.8 to 1682.3mg m-2 h-1. CO2 fluxes from mangroves studied in southern China have ranged from 0 to 904.6mg m−2 h−1 [22-26], Can Gio mangrove in Vietnam presented CO2 fluxes from 234.1 to 953.3mg m−2 h−1 [27] and New Caledonia 57.4 to 389.4mg m−2 h−1 [28-30].
The highest fluxes of CO2 (1209.3 ± 297.1mg m-2 h-1) in this study were recorded in the dry and warm seasons, under conditions of low humidity and high soil temperature (Table 2). The lowest fluxes (459.0 ± 154.2mg m-2 h-1) were in nortes season (Figure 2A). A high CO2 flux to the atmosphere with warmer temperatures was consistent with other studies in mangroves [31,32]. A high temperature in the mangrove soil led to an increase in microbial activity which generates higher levels of soil respiration and CO2 emissions [33,34]. Also, the rise in temperature accelerates the decomposition of organic matter and consequently produces and emits CO2 [35].
During the dry and warm seasons, the soils were never flooded and according to Chen et al. [22] and Alongi [36], CO2 emission rates from mangrove soil tend to be higher when soils are exposed to the atmosphere, because molecular diffusion is faster for gases than for fluids, increasing the aerobic respiration and accelerating chemical oxidation of soil organic matter. When the temperature decreased during the nortes season, CO2 emissions from mangrove soils also dropped. During the rainy season, low CO2 fluxes were observed despite the temperatures were around 26.6 to 27.2°C, so precipitation may influence the fluxes [27]. In flooded soils, anoxic conditions can be induced by limiting the decomposition processes of organic matter, hence CO2 production diminishes [37].


Methane fluxes
CH4 fluxes ranged between 592.8 to 8909μg m−2 h−1 which were lower than those recorded in the mangroves of Shenzhen and Hong Kong in South China ranging from 0 to 82916μg m−2 h−1 [22-24]. The measured fluxes were similar to those recorded at the Bhitarkanika mangrove in India (2300μg m−2 h−1) [38], Taiwan (5071.1μg m−2 h−1) [39] and Jiulong River, China (5360.1μg m−2 h−1) [24], but higher than some mangroves in New Caledonia, Brazil, and Indonesia [28,40,41].
The CH4 fluxes were significantly higher in the rainy (4957.2 ± 1478.7µg m-2 h-1) and nortes seasons (3688 ± 2325.3µg m-2 h-1), and lower fluxes were observed during the dry season (1278 ± 393.9µg m-2 h-1) (Figure. 2B). The CH4 dynamics recorded in the Terminos lagoon was like other studies that attribute the variation of CH4 emissions to sediment temperature and water level [42].
According to Chauhan et al. [38] sediment temperature is responsible for the seasonal variation of CH4 emission. By decreasing the temperature, the decomposition of organic matter is reduced, producing methanogens. Methanogenic bacteria are responsible for releasing CH4 and are active anaerobes in flooded and swampy areas [43,44]. It has been reported that the microbiological process that generates the formation of methane in the soil is controlled by edaphic factors among them, the redox potential (Eh) and the pH of the soil. The Eh during the rainy and northern seasons were from -105 to -205mV in the upper layer (0-30cm depth) and from -115 to -226 in the 30-60cm in the bottom layer (Table 2). A redox potential of -150 to -200mV favors methanogenesis in the natural wetlands which might have resulted in higher fluxes of methane [45]. pH ranged from 6.2 to 7.8 and are considered optimal for methane production [46].
High organic matter content and low Eh indicate that the degradation of more organic matter content induces greater CH4 production, while high Eh indicates that the soil has adequate ventilation, conducting soil respiration and CO2 emission [33].
Nitrogen oxid
N2O fluxes varied significantly between sampling seasons (375.4 to 1002.2µg m-2 h-1) but were in the range of reports from other authors [47,48]. The N2O flux was higher in the rainy season (772.9 ± 138.5µg m-2 h-1) and lower in nortes season (464.2 ± 65.4µg m-2 h-1; Figure 2C). The seasonal variation of N2O is like that recorded by Allen et al. [49] who identified that sediment temperature and moisture are the key to seasonal differences in N2O fluxes.
Global warming potential
The CO2-eq emission rate was 86.9 ± 31.7 Mg CO2-eq ha-1 yr-1 with a contribution of 71.6% CO2, 10.3% CH4, and 18.1% N2O. It had significant differences between seasons (p < 0.001). The CO2-eq was higher in the dry season (Figure 3). There was also a significant difference between sites (p = 0.01), the ATA and EP mangroves contributed with 26.2% and 23.4% of the CO2-eq respectively. The highest contribution to global warming potential among season and sites was CO2 (62.2 ± 34.1 Mg CO2-eq ha-1 yr-1) followed by N2O (15.7 ± 4.5 Mg CO2-eq ha-1 yr-1) and CH4 (9 ± 5.7 Mg CO2-eq ha-1 yr-1). The CO2-eq contribution has been observed in other studies [50-52]. Allen et al. [53] mention that in subtropical mangrove sediments during low CH4 emissions, the N2O has a high global warming potential that can dominate trace gas emissions, that pattern coincides with our results (Figure 3).

Regardless of the cause, the loss of mangroves leads to a decrease in the organic carbon inventory, especially if the soil horizon is disturbed, which can be converted into CO2-eq emissions to the atmosphere [54]. The CO2-eq observed in the degraded mangroves (NCD and TOR) was 70.3 ± 22.8 Mg CO2-eq ha-1 yr-1, where the contribution of CO2 was 56%, N2O 23.8% and CH4 19.8%; Thus, in degradation conditions, the emission of N2O and CH4 increases and that of CO2 decreases.
GHG fluxes in conserved and degraded mangroves
The ecological condition of each mangrove affects the GHG fluxes. CO2 and CH4 fluxes had significant differences (p < 0.05) between conserved and degraded mangroves. ATA had the highest flux of CO2 (966.4 ± 528.1mg m-2 h-1), followed by the EP (859.8 ± 393.0mg m-2 h-1). TOR had the lowest CO2 fluxes (206.8 ± 24.9mg m-2 h-1) (Figure 2A). The highest CH4 fluxes were measured in NCD (4736.1 ± 2588.4µg m-2 h-1) and TOR (6354 ± 453.7µg m-2 h-1) which have an important degradation. N2O did not show significant differences (p > 0.05), ATA had 713.6 ± 258.1µg m-2 h-1, NCD 763.9 ± 219.6µg m-2 h-1 and NCC 634 ± 194.7µg m-2 h-1. So, the highest CO2 fluxes were measured in conserved mangroves, and the lower in degraded mangroves. The state of the mangrove (conserved or degraded) is observed in the PCA of Figure 4. The first two axes explained 72.5% of the variance, in PC1 the factors with the greatest contribution were NO3-, soil organic matter (SOM) and Eh. In PC2 were EC, Ts, and SO42-. The sediments presented groups related to GHG emissions. That is, the PCA showed that the conserved mangroves were grouped because they have high values of CO2, N2O, Ts, EC, Eh, NO3-, and SOM. The degraded mangroves had relevant values of CH4, pH, and SM, and poor values of SO42-.
At ATA, EP, and NCC the concentration of SO42- ranged between 16 and 36mg kg-1 and the NO3- between 30 and 66mg kg-1. The SOM registered values between 9.3 - 16.4% (Table 2). Rhizophora mangle and Laguncularia racemosa dominate in ATA and EP and have a large leaf litter cover which is essential in the nutrient cycle, since its production and decomposition represents the main transfer of organic matter and nutrients from the aerial part to the surface of the soil [55,56]. Soils with litter produce carbon-rich compounds that are more susceptible to bacterial decomposition [57]. Microorganisms promote the decomposition of organic matter by nitrogen fixation, the reduction of sulfates and the production of enzymes [58]. In NCC, Avicennia germinans dominates the area, which has pneumatophores that allow the exchange of oxygen and CO2 from the root system of the plant. Troxler et al. [59] mentions that black mangrove pneumatophores also contribute more average flux of CO2 from the soil to the atmosphere.
Soil microorganisms, respiration of plant root cells and the degradation of leaf litter produce CO2 [60]. The emission of CO2 to the atmosphere occurs when microorganisms oxidize organic C using O2, NO3-, Mn4+, Fe3+, and SO42- as electron acceptors [40,61]. The correlation of SOM, nutrients and high CO2 emissions is related to site-specific conditions and biotic and abiotic factors. Hien et al. [35] mention that the development of mangroves and the enrichment of organic matter in the sediments also influences the properties of the sediment, modifying the production and emission of CO2. Mangrove disturbance can also alter natural soil SOM decomposition and storage processes. When mangroves degrade the SOM can become mineralized, contributing to more CO2 emissions [62].
NCD and TOR had the highest CH4 fluxes. These sites show a negative correlation between CH4 and Eh, SO42-, and EC, and positive with pH (Figure 4). High values of salinity and SO42- favor the activity of sulfate-reducing bacteria and hinder the metabolism of methanogens, since they inhibit the production of CH4 from wetlands [63,64]. Methanogens are anaerobic and for methanogenesis to occur the redox potential must be as low as -200mV, and other competing terminal electron acceptors must have been reduced (O2, NO3-, and SO42-) [65]. Previous studies have found that the CH4 potential soil production and anaerobic and aerobic oxidation have a negative correlation with soil redox potential [66] and with high salinity concentrations [38,45]. On the other hand, a pH of 7.6 facilitates the growth of methanogens and the production of CH4 [38]. Other authors have also reported that in sites with loss of vegetation the flux of CH4 increases considerably compared to pristine sites [50].
Soil N2O flux did not present a clear pattern between conserved and degraded mangroves and no significant differences were observed (p = 0.149). ATA and NCC had the highest N2O fluxes in the dry season and NCD the highest in the rainy season (Figure 2C). ATA and NCC had positive correlations of N2O with Eh, NO3-, and EC; while NCD recorded negative correlations of N2O with Eh, NO3-, and EC (Figure 4). Eh was positive in dry conditions and negative in rains and nortes; so, changes in the hydrological regime influence soil redox conditions and can alter nitrogen availability [67,68].
Nitrogen oxide emissions from soils are related to microbial processes of nitrification (oxidation of NH4+ to NO2- and NO3-) and denitrification (reduction of NO3- and NO2- to NO, N2O and N2) [69]. Both processes use soil mineral nitrogen (NH4+ and NO3-) that originate mainly from the mineralization of organic matter or fertilization.
In dry conditions, denitrification under aerobic environments carried out by chemolithotrophic nitrifying bacteria with positive Eh oxidizes NH4+ to NO3- and dominates the production of N2O [70]. While in rains and nortes, under anaerobic conditions, the sediment had high levels of nitrates and negative Eh in the conserved sites (Table 2 & 3) that allows denitrification.

Soil carbon stock
The mean SCS was not significantly different between climatic periods (p = 0.88) and between sampling depths (p = 0.35). The total soil carbon stock (0 - 60cm depth) was 431 ± 180.9 Mg C ha-1 during the nortes, 419 ± 142.8 Mg C ha-1 on the rainy and 415 ± 103.6 Mg C ha-1 on the dry seasons. In the upper layer (0-30cm), the mean was 201± 71.8 Mg C ha-1, and for the 30-60cm layer was 221 ± 70.3 Mg C ha-1 (Figure 5). The carbon stored in mangrove soil is consistent with other studies carried out in Laguna de Terminos [71] and other places of Mexico [72], as well as other parts of the world [73], but higher than Sofala Bay [74]; Dominican Republic [75]; Florida [76] and Malaysia [77] (Table 4). Zamora et al. [12] quantified the carbon reservoir in Mexican coastal forested wetlands from 79 - 650 Mg C ha-1. The observed differences are probably related to the measured soil depth.

The ATA stores 591.3 ± 85.1, EP 516.3 ± 53, NCC 403.4 ± 23.5, NCD 287.9 ± 22.4 and TOR 255.8 ± 16.4 Mg C ha-1 reflecting different conditions of ecological degradation.
Soils showed an apparent density ranged between 0.7 - 1.1g cm-3 in the most superficial layer and 1.0 - 1.3g cm-3 in the 30 - 60cm layer (Table 2). Mitsch & Gosselink [65] mention that mineral soils have apparent densities ranged on 1 - 2g cm-3, so it is likely that some horizons correspond to organic soils and the deeper layer to mineral soils.
The preserved mangroves had the highest SOM and SOC. The highest concentrations were observed in the rainy season (between 0 - 30cm) and nortes throughout the sediment column (0 - 60cm, Table 2). Those seasons had intense rains and tides that flooded the sites, the soil generates reserves of organic carbon due to the stagnant water for long periods, delaying the oxidation of SOM and increasing the accumulation rate of SOC due to the anaerobic conditions or a lower soil temperature relative to the ambient [78]. In the dry season, the low water content and CO2 concentration of the soil reduce the stored carbon intensifying the flux of CO2 [79].



The percentage of sand ranged between 75 and 89%, clay 5 to 10% and silt 6 to 16%, fine sediments (clay + silt) were greater in conserved than degraded mangroves (Figure 6) and together with the sediment humidity influence the SCS, since the water content and the percentage of fine sediments have a positive impact on the surface SOC [80,81]. On the other hand, Schile et al. [82] mentions that the texture of the sediment is one of the factors that regulates the preservation of organic matter, which explains why carbon reserves are poor in sandy soils.
The results suggest that carbon storage is influenced by the ecological conditions of the sites that induce changes in soil properties such as moisture, texture, the amount of SOM, SOC, bulk density and soil depth (Figure 4).
Degraded sites have lower SCS in the 0 - 30cm layer of the sediment (Figure 5). According to Lovelock et al. [83] the disturbance and exposure of sediment in degraded mangroves accelerates the erosion of the upper layer, leaving it exposed to physical forces that lead to chemical weathering and higher decomposition rates, which leads to lower carbon storage. Senger et al. [10] found that carbon is rich in the upper 30cm of the sediment in intact sites and is poor in degraded sites. Likewise, intact plots have more SOC associated with higher CO2 fluxes, while degraded plots have a higher SOC low and poorer CO2 fluxes, which is like our results.
Degradation of mangroves leads to higher CO2 emission and changes the status from carbon sink to carbon source [84] which significantly contributes to global warming and climate change.
Conclusion
GHG fluxes were directly related to seasonal variation and to environmental factors specific to each mangrove. Seasonality influenced soil conditions; flooded conditions favored CH4 and N2O production, while the dry season with more oxygenated soils favored CO2 production. CO2 and N2O fluxes were higher in conserved sites controlled by redox potential, soil temperature and electrical conductivity, while CH4 fluxes were higher in degraded mangroves controlled by pH, SO4 and soil moisture. The highest carbon storage was observed in conserved mangroves. Ecological degradation in mangroves can increase GHG emissions and reduce the ecosystem's ability to store carbon. Therefore, mangrove conservation and restoration are essential to mitigate GHG emissions and maintain their function as carbon sinks.
Acknowledgement
The authors thank Faustino Zavala Garcia and María Isabel Saavedra for their assistance with laboratory analyses, graphing and reviewing the manuscript.
References
- Donato DC, Kauffman JB, Murdiyarso D, Kurnianto S, Stidham M, et al. (2011) Mangroves among the most carbon-rich forests in the tropics. Nature Geoscience 4(5): 293-297.
- Himes-Cornell A, Grose SO, Pendleton L (2018) Mangrove ecosystem service values and methodological approaches to valuation: where do we stand? Frontiers in Marine Science 5: 376.
- Lee SY, Primavera JH, Dahdouh‐Guebas F, McKee K, Bosire JO, et al. (2014) Ecological role and services of tropical mangrove ecosystems: a reassessment. Global Ecology and Biogeography 23(7): 726-743.
- Kauffman JB, Adame MF, Arifanti VB, Schile‐Beers LM, Bernardino AF, et al. (2020) Total ecosystem carbon stocks of mangroves across broad global environmental and physical gradients. Ecological Monographs 90(2): e01405.
- Hamilton SE, Friess DA (2018) Global carbon stocks and potential emissions due to mangrove deforestation from 2000 to 2012. Nature Climate Change 8(3): 240-244.
- Kauffman JB, Hernandez Trejo H, del Carmen Jesus Garcia M, Heider C, Contreras WM (2016) Carbon stocks of mangroves and losses arising from their conversion to cattle pastures in the Pantanos de Centla, Mexico. Wetlands Ecology and Management 24(2): 203-216.
- WWF (World Wildlife Federation) (2020). In: Almond REA, Grooten M, Petersen T (Eds.), Living Planet Report 2020 - Bending the Curve of Biodiversity Loss. WWF, Gland, Switzerland.
- Osland MJ, Feher LC, Griffith KT, Cavanaugh KC, Enwright NM, et al. (2017) Climatic controls on the global distribution, abundance, and species richness of mangrove forests. Ecological Monographs 87(2): 341-359.
- Lovelock CE, Fourqurean JW, Morris JT (2017) Modeled CO2 emissions from coastal wetland transitions to other land uses: tidal marshes, mangrove forests, and seagrass beds. Frontiers in Marine Science 4: 143.
- Senger DF, Hortua DS, Engel S, Schnurawa M, Moosdorf N, et al. (2021) Impacts of wetland dieback on carbon dynamics: A comparison between intact and degraded mangroves. Science of the Total Environment 753: 141817.
- Oertel C, Matschullat J, Zurba K, Zimmermann F, Erasmi S (2016) Greenhouse gas emissions from soils—A review. Geochemistry 76(3): 327-352.
- Zamora S, Sandoval-Herazo LC, Ballut-Dajud G, Del Ángel-Coronel OA, Betanzo-Torres EA, et al. (2020) Carbon fluxes and stocks by Mexican tropical forested wetland soils: a critical review of its role for climate change mitigation. International Journal of Environmental Research and Public Health 17(20): 7372.
- Adame MF, Zakaria RM, Fry B, Chong VC, Then YHA, et al. (2018) Loss and recovery of carbon and nitrogen after mangrove clearing. Ocean & Coastal Management 161: 117-126.
- Yáñez-Arancibia A, Twilley RR, Lara-Domínguez AL (1998) Los ecosistemas de manglar frente al cambio climático global. Madera y bosques 4(2): 3-19.
- Guerra-Santos JJ, Kahl JD (2018) Redefining the seasons in the Términos Lagoon region of Southeastern México: May is a transition month, not a dry month. Journal of Coastal Research 34(1): 193-201.
- Howard J, hoyt S, Isensee K, Telszewski M, Pidgeon E (Eds.) (2014) Coastal blue Carbon: Methods for assessing carbon stocks and emissions factors in mangroves, tidal salt marshes, and seagrasses. Conservation International, Intergovernmental Oceanographic Commission of UNESCO, International Union for Conservation of Nature. Arlington, Virginia, USA.
- IPCC, 2021: Summary for policymakers. in: Climate change 2021: The physical science basis. Contribution of working group i to the sixth assessment report of the intergovernmental panel on climate change.
- Etchevers B, Jorge D (2005) Manual para la determinación de carbono en la parte aérea y subterránea de sistemas de producción en laderas.
- Gee GW, Bauder JW (1986) Particle‐size analysis. Methods of soil analysis: Part 1 Physical and Mineralogical Methods 5: 383-411.
- Heiri O, Lotter AF, Lemcke G (2001) Loss on ignition as a method for estimating organic and carbonate content in sediments: reproducibility and comparability of results. Journal of Paleolimnology 25: 101-110.
- Kauffman JB, Donato DC (2012) Protocols for the measurement, monitoring and reporting of structure, biomass and carbon stocks in mangrove forests. Working Paper 86. CIFOR, Bogor, Indonesia.
- Chen GC, Tam NFY, Ye Y (2010) Summer fluxes of atmospheric greenhouse gases N2O, CH4 and CO2 from mangrove soil in South China. Science of the Total Environment 408(13): 2761-2767.
- Wang T, Zhang H, Lin H, Fang C (2015) Textural–spectral feature-based species classification of mangroves in Mai Po Nature Reserve from Worldview-3 imagery. Remote Sensing 8(1): 24.
- Chen G, Chen B, Yu D, Tam NF, Ye Y, et al. (2016) Soil greenhouse gas emissions reduce the contribution of mangrove plants to the atmospheric cooling effect. Environmental Research Letters 11(12): 124019.
- Zheng X, Guo J, Song W, Feng J, Lin G (2018) Methane emission from mangrove wetland soils is marginal but can be stimulated significantly by anthropogenic activities. Forests 9(12): 738.
- Xu Y, Liao B, Jiang Z, Xin K, Xiong Y, et al. (2021) Emission of greenhouse gases (CH4 and CO2) into the atmosphere from restored mangrove soil in South China. Journal of Coastal Research 37(1): 52-58.
- Van Vinh T, Allenbach M, Joanne A, Marchand C (2019) Seasonal variability of CO2 fluxes at different interfaces and vertical CO2 concentration profiles within a Rhizophora mangrove forest (Can Gio, Viet Nam). Atmospheric Environment 201: 301-309.
- Leopold A, Marchand C, Deborde J, Chaduteau C, Allenbach M (2013) Influence of mangrove zonation on CO2 fluxes at the sediment–air interface (New Caledonia). Geoderma 202: 62-70.
- Leopold A, Marchand C, Deborde J, Allenbach M (2015) Temporal variability of CO2 fluxes at the sediment-air interface in mangroves (New Caledonia). Science of the Total Environment 502: 617-626.
- Jacotot A, Marchand C, Allenbach M (2019) Increase in growth and alteration of C: N ratios of Avicennia marina and Rhizophora stylosa subject to elevated CO2 concentrations and longer tidal flooding duration. Frontiers in Ecology and Evolution 7: 98.
- Chen GC, Tam NF, Ye Y (2012) Spatial and seasonal variations of atmospheric N2O and CO2 fluxes from a subtropical mangrove swamp and their relationships with soil characteristics. Soil Biology and Biochemistry 48: 175-181.
- Gnanamoorthy P, Selvam V, Burman PKD, Chakraborty S, Karipot A, et al. (2020) Seasonal variations of net ecosystem (CO2) exchange in the Indian tropical mangrove forest of Pichavaram. Estuarine, Coastal and Shelf Science 243.
- Poungparn S, Komiyama A, Tanaka A, Sangtiean T, Maknual C, et al. (2009) Carbon dioxide emission through soil respiration in a secondary mangrove forest of eastern Thailand. Journal of Tropical Ecology 25(4): 393-400.
- Simpson LT, Osborne TZ, Feller IC (2019) Wetland soil CO2 efflux along a latitudinal gradient of spatial and temporal complexity. Estuaries and Coasts 42: 45-54.
- Hien HT, Marchand C, Aimé J, Cuc NTK (2018) Seasonal variability of CO2 emissions from sediments in planted mangroves (Northern Viet Nam). Estuarine, Coastal and Shelf Science 213: 28-39.
- Alongi D (2009) The energetics of mangrove forests. Springer Science & Business Media. Springer, ISBN 978-1-4020-4270-6.
- Kristensen E, Connolly RM, Otero XL, Marchand C, Ferreira TO, et al. (2017) Biogeochemical Cycles: Global Approaches and Perspectives. In: Rivera-Monroy V, Lee S, Kristensen E, Twilley R (Eds.), Mangrove Ecosystems: A Global Biogeographic Perspective. Springer, Cham, pp. 163-209.
- Chauhan R, Datta A, Ramanathan AL, Adhya TK (2015) Factors influencing spatio-temporal variation of methane and nitrous oxide emission from a tropical mangrove of eastern coast of India. Atmospheric Environment 107: 95-106.
- Lin CW, Kao YC, Chou MC, Wu HH, Ho CW, et al. (2020) Methane emissions from subtropical and tropical mangrove ecosystems in Taiwan. Forests 11(4): 470.
- Nóbrega GN, Ferreira TO, Neto MS, Queiroz HM, Artur AG, et al. (2016). Edaphic factors controlling summer (rainy season) greenhouse gas emissions (CO2 and CH4) from semiarid mangrove soils (NE-Brazil). Science of the Total Environment 542: 685-693.
- Chen GC, Ulumuddin YI, Pramudji S, Chen SY, Chen B, et al. (2014) Rich soil carbon and nitrogen but low atmospheric greenhouse gas fluxes from North Sulawesi mangrove swamps in Indonesia. Science of the Total Environment 487: 91-96.
- Lekphet S, Nitisoravut S, Adsavakulchai S (2005) Estimating methane emissions from mangrove area in Ranong Province, Thailand. Songklanakarin Journal of Science and Technology 27(1): 153-163.
- Whalen, SC (2005) Biogeochemistry of methane exchange between natural wetlands and the atmosphere. Environmental Engineering Science 22(1): 73-94.
- Pazinato JM, Paulo EN, Mendes LW, Vazoller RF, Tsai SM (2010) Molecular characterization of the archaeal community in an Amazonian wetland soil and culture-dependent isolation of methanogenic archaea. Diversity 2(7): 1026-1047.
- Olsson L, Ye S, Yu X, Wei M, Krauss KW, et al. (2015) Factors influencing CO2 and CH4 emissions from coastal wetlands in the Liaohe Delta, Northeast China. Biogeosciences 12(16): 4965-4977.
- Oremland RS (1988) Biogeochemistry of methanogenic bacteria. In: AJB Zehnder, WS Tumm (Eds.), Biology of Anaerobic Microorganisms. John Wiley, New York, pp. 641-705.
- Konnerup D, Betancourt-Portela JM, Villamil C, Parra JP (2014) Nitrous oxide and methane emissions from the restored mangrove ecosystem of the Ciénaga Grande de Santa Marta, Colombia. Estuarine, Coastal and Shelf Science 140: 43-51.
- Huang Y, Wang C, Lin C, Zhang Y, Chen X, et al. (2019) Methane and nitrous oxide flux after biochar application in subtropical acidic paddy soils under tobacco-rice rotation. Scientific Reports 9(1): 17277.
- Allen D, Dalal RC, Rennenberg H, Schmidt S (2011) Seasonal variation in nitrous oxide and methane emissions from subtropical estuary and coastal mangrove sediments, Australia. Plant Biology 13(1): 126-133.
- Romero-Uribe HM, López-Portillo J, Reverchon F, Hernández ME (2022) Effect of degradation of a black mangrove forest on seasonal greenhouse gas emissions. Environmental Science and Pollution Research 29(8): 11951-11965.
- Cameron C, Hutley LB, Munksgaard NC, Phan S, Aung T, et al. (2021) Impact of an extreme monsoon on CO2 and CH4 fluxes from mangrove soils of the Ayeyarwady Delta, Myanmar. Science of the Total Environment 760: 143422.
- Cameron C, Hutley LB, Friess DA, Munksgaard NC (2019) Hydroperiod, soil moisture and bioturbation are critical drivers of greenhouse gas fluxes and vary as a function of land use change in mangroves of Sulawesi, Indonesia. Science of the Total Environment 654: 365-377.
- Allen DE, Dalal RC, Rennenberg H, Meyer RL, Reeves S, et al. (2007) Spatial and temporal variation of nitrous oxide and methane flux between subtropical mangrove sediments and the atmosphere. Soil Biology and Biochemistry 39(2): 622-631.
- Alongi DM (2020) Carbon cycling in the world’s mangrove ecosystems revisited: Significance of non-steady state diagenesis and subsurface linkages between the forest floor and the coastal ocean. Forests 11(9): 977.
- Isaac SR, Nair MA, Suresh PR (2006) Biological degradation of Mahogany (Swietenia macrophylla) leaf litter. Journal of the Indian Society of Soil Science 54(1): 117-119.
- Reef R, Feller IC, Lovelock Ce (2010) Nutrition of mangroves. Tree Physiology 30(9): 1148-1160.
- Friesen SD, Dunn C, Freeman C (2018) Decomposition as a regulator of carbon accretion in mangroves: a review. Ecological Engineering 114: 173-178.
- Sahoo K, Dhal NK (2009) Potential microbial diversity in mangrove ecosystems: a review. Indian Journal of Marine Sciences 38(2): 249-256.
- Troxler TG, Barr JG, Fuentes JD, Engel V, Anderson G, et al. (2015) Component-specific dynamics of riverine mangrove CO2 efflux in the Florida coastal Everglades. Agricultural and Forest Meteorology 213: 273-282.
- Kristensen E, Bouillon S, Dittmar T, Marchand C (2008) Organic carbon dynamics in mangrove ecosystems: a review. Aquatic Botany 89(2): 201-219.
- Luo M, Huang JF, Zhu WF, Tong C (2019). Impacts of increasing salinity and inundation on rates and pathways of organic carbon mineralization in tidal wetlands: a review. Hydrobiologia 827: 31-49.
- Arias-Ortiz A, Masqué P, Glass L, Benson L, Kennedy H, et al. (2021) Losses of soil organic carbon with deforestation in mangroves of Madagascar. Ecosystems 24: 1-19.
- Vizza C, West WE, Jones SE, Hart JA, Lamberti GA (2017) Regulators of coastal wetland methane production and responses to simulated global change. Biogeosciences 14(2): 431-446.
- Yang P, Wang MH, Lai DY, Chun KP, Huang JF, et al. (2019) Methane dynamics in an estuarine brackish Cyperus malaccensis marsh: Production and porewater concentration in soils, and net emissions to the atmosphere over five years. Geoderma 337: 132-142.
- Mitsch WJ, Gosselink JG (2007) Wetlands, John Wiley & Sons, Inc., Hoboken, New Jersey, USA.
- Wang W, Zeng C, Sardans J, Wang C, Tong C, et al. (2018) Soil methane production, anaerobic and aerobic oxidation in porewater of wetland soils of the Minjiang River Estuarine, China. Wetlands 38: 627-640.
- Verhoeven J (2009) Wetland Biogeochemical Cycles and their Interactions. In: E Maltby, T Barker (Eds.), The Wetlands Handbook, 266. Blackwell Publishing, ISBN 978-0-632-05255-4.
- Verhoeven JT, Laanbroek HJ, Rains MC, Whigham DF (2014) Effects of increased summer flooding on nitrogen dynamics in impounded mangroves. Journal of Environmental Management 139: 217-226.
- Butterbach-Bahl K, Baggs EM, Dannenmann M, Kiese R, Zechmeister-Boltenstern S (2013) Nitrous oxide emissions from soils: how well do we understand the processes and their controls? Philosophical Transactions of the Royal Society B: Biological Sciences 368(1621).
- Bauza JF, Morell JM, Corredor, JE (2002) Biogeochemistry of nitrous oxide production in the red mangrove (Rhizophora mangle) forest sediments. Estuarine, Coastal and Shelf Science 55(5): 697-704.
- Moreno-May G, Cerón J, Cerón R, Guerra J, Amador L, et al. (2010) Evaluation of carbon storage potential in mangrove soils of Isla del Carmen. Unacar Tecociencia 4: 23-39.
- Hernández ME, Junca-Gómez D (2020) Carbon stocks and greenhouse gas emissions (CH4 and N2O) in mangroves with different vegetation assemblies in the central coastal plain of Veracruz Mexico. Sci Total 741: 140276.
- Savari A, Khaleghi M, Safahieh AR, Hamidian Pour M, Ghaemmaghami S (2020) Estimation of biomass, carbon stocks and soil sequestration of Gowatr mangrove forests, Gulf of Oman. Iranian Journal of Fisheries Sciences 19(4): 1657-1680.
- Sitoe AA, Comissario Mandlate LJ, Guedes BS (2014) Biomass and carbon stocks of Sofala Bay mangrove forests. Forests 5(8): 1967-1981.
- Kauffman JB, Heider C, Norfolk J, Payton F (2014) Carbon stocks of intact mangroves and carbon emissions arising from their conversion in the Dominican Republic. Ecological Applications 24(3): 518-527.
- Doughty CL, Langley JA, Walker WS, Feller IC, Schaub R, et al. (2016) Mangrove range expansion rapidly increases coastal wetland carbon storage. Estuaries and Coasts 39: 385-396.
- Besar NA, Suhaili NS, Fei JLJ, Shaari FW, Idris MI, et al. (2020). Carbon stock estimation of mangrove forest in sulaman lake forest reserve, Sabah, Malaysia. Biodiversitas Journal of Biological Diversity 21(12).
- Dayathilake DDTL, Lokupitiya E, Wijeratne VPIS (2021) Estimation of soil carbon stocks of urban freshwater wetlands in the Colombo Ramsar Wetland City and their potential role in climate change mitigation. Wetlands 41(2): 29.
- Maier M, Schack-Kirchner H, Hildebrand EE, Schindler D (2011) Soil CO2 efflux vs. soil respiration: Implications for flux models. Agricultural and Forest Meteorology 151(12): 1723-1730.
- Ouyang X, Lee SY, Connolly RM (2017) Structural equation modelling reveals factors regulating surface sediment organic carbon content and CO2 efflux in a subtropical mangrove. Science of the Total Environment 578: 513-522.
- Xiong Y, Liao B, Proffitt E, Guan W, Sun Y, et al. (2018) Soil carbon storage in mangroves is primarily controlled by soil properties: A study at Dongzhai Bay, China. Science of the Total Environment 619: 1226-1235.
- Schile LM, Kauffman JB, Crooks S, Fourqurean JW, Glavan J, et al. (2017). Limits on carbon sequestration in arid blue carbon ecosystems. Ecological Applications 27(3): 859-874.
- Lovelock CE, Ruess RW, Feller IC (2011) CO2 efflux from cleared mangrove peat. PloS one 6(6): e21279.
- Alongi DM (2015) The impact of climate change on mangrove forests. Current Climate Change Reports 1: 30-39.















